MICRO-BODIES
Both plant and animal cells contain very small granular bead like structures in their protoplasm, surrounded by a single unit membrane. Such structures are different from lysosomal vesicles because of their specific physiological properties; they are called micro bodies. Their size varies from 0.6mm – 1.5mm. The number also varies from 20 – 100 per cell. In germinating seeds micro bodies increase in their number considerably. The contents of such structures may be amorphous, crystalline or fibrillar in nature. However, the enzymatic contents in them vary. Based on their functional properties they are classified into peroxisomes and glyoxysomes.
Peroxisomes
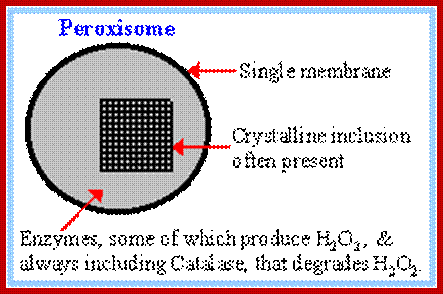
Peroxisomes are characterized by their peroxidase activity. They are more or less dense spherical bodies bounded by a single unit membrane. The size is about 0.6mm – 0.8mm. Almost every species in the plant kingdom contain peroxisomes in most of the cells. But among animals only higher vertebrates contain such structures. In higher plants, leaves of the C4 plants also contain micro bodies. They are mostly associated with plastids and mitochondria and they are mainly responsible for photo-respiration.
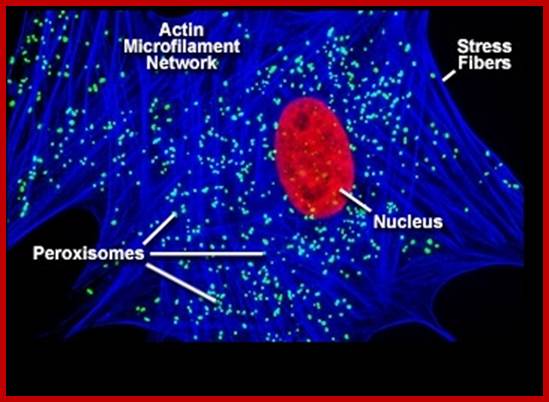
Fine dots are peroxisome.www.pixgood.com; http://flipper.diff.org/
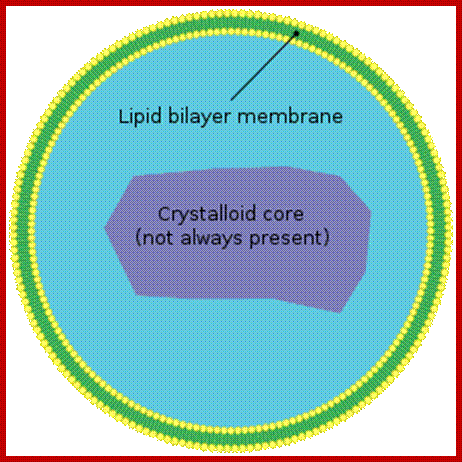
Basic structural Feature of Peroxisome; It is enveloped by single unit membrane and contains crystal structure inside. https://en.wikipedia.org
Enzyme contents
The most important enzyme component of this cell organelle is peroxidase or catalase or both. It also contains glycollate oxidase, glutamate glyoxylase, transaminases, other enzymes required for the breakdown of fatty acids. The co-enzymes present are FAD, NAD and NADP. At least there are 32 known peroxisome proteins.
Peroxisomes assemble from derivative of ER vesicles and they replicate by fission. Peroxisomal proteins translated contain specific peroxisomal targeting sequences PTS at the C-end of proteins (PTS1) or at N-terminal called PTS2. They are imported in unfolded condition. Protein receptors PLX5 and PEX7 with cargo proteins contain PTS1 and PTS2 amino acid sequences. Once the proteins are delivered, they return to cytoplasm. By what is called recycling mechanism or shuttle mechanism for which they require ATP input.
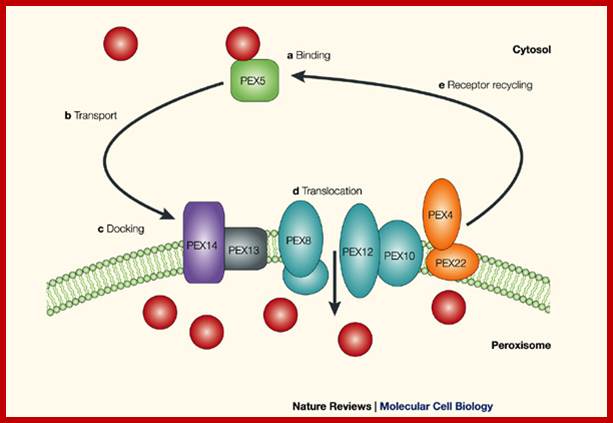
Peroxisomal protein import; www.Nature.com
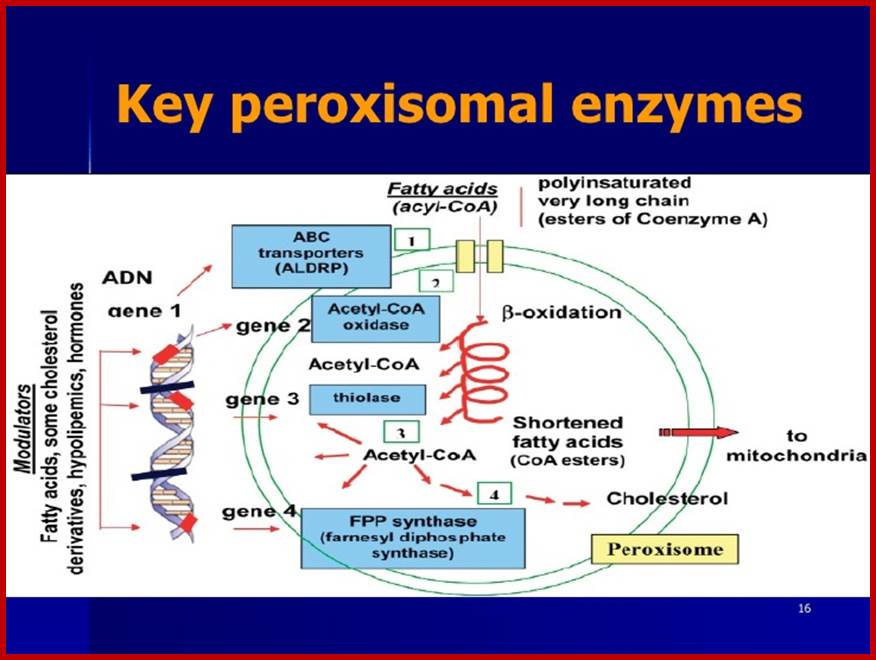
Key peoxisomal enzymes; www.slideshare.net
FUNCTIONS:-
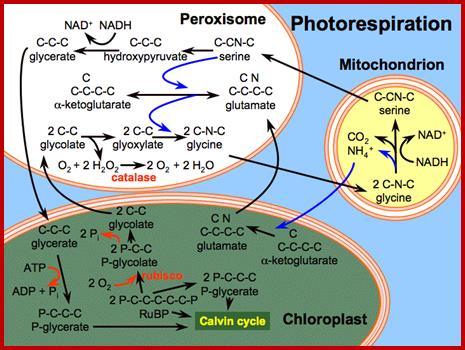
In C3 plants like dicots, peroxisomes are mainly involved in photorespiration. When the intracellular concentration of oxygen is high; RUBP carboxylase present in chloroplasts acts as oxygenase and splits ribulose diphosphate into phosphoglycerate and phospho glycollate. The latter is transported into peroxisomes, where phospho glycollate is dephosphorylatated and converted to glyoxalate. Then it is converted to Glycine in turn converted to serine in mitochondria by decarboxylation process. As oxygen is utilized this process it is called photorespiration which is insensitive to cyanide; thus it is distinct from respiratory oxidation.
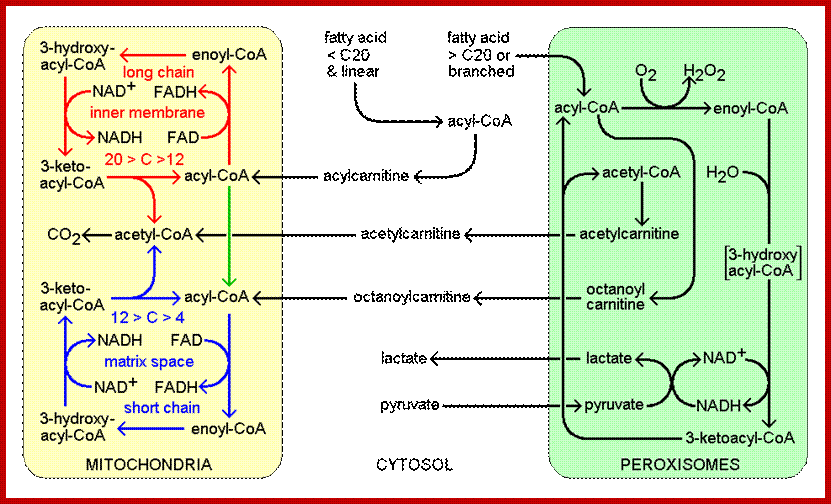
http://flipper.diff.org/
Biogenesis; Pulse labeling and chase experiments with specific substrates reveal that the peroxisomal proteins are synthesized free from ER membranes. Then they are transported to peroxisomes or glyoxysomes directly through specific carriers which use specific peroxisomal or glyoxysomal protein signature sequences and deposit on to their vesicle membrane translocators. The life span of peroxisomes is about 45 days.
Glyoxysomes
Glyoxysomes are distinct from peroxisomes because of different functions. They are found in fungi and other higher plants. Storage organs rich in oil globules contain greater number of glyoxysomes. Such membranous structures loaded with oil globules are also called spherosomes of 0.5 to 1.5 nm size and they are associated with mitochondria. When such oil laden seeds germinate, the number of glyoxysomes increases significantly and remains very active.
Glyoxysomes contain a host of enzymes responsible for b-oxidation of a fatty acids, citrate synthase, isocitrate lyase, malate synthase, hydrogenase, urate oxidase, etc. Thus it has all the enzymes necessary for fatty acid oxidation as well as glyoxylate cycle and gluconeogenesis.
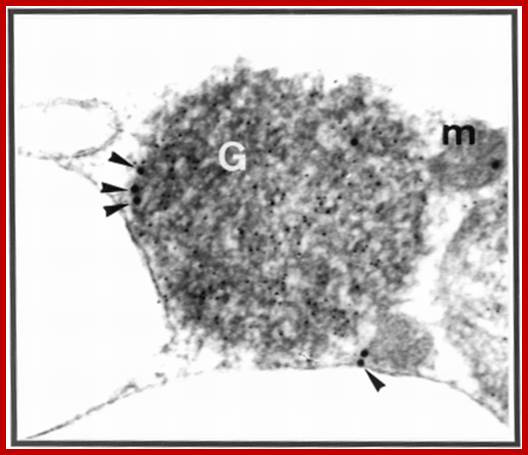
A representative electron micrograph of cucumber cotyledon thin sections illustrating results of indirect, double immunogold labeling with potato porin antiserum and anti-pumpkin seed catalase. The 15-nm gold particles show the peripheral localization of porin. The 5-nm gold particles show the matrix localization of catalase. G, glyoxysome; m, mitochondrion. The magnification is 90,000×. http://pcp.oxfordjournals.org/
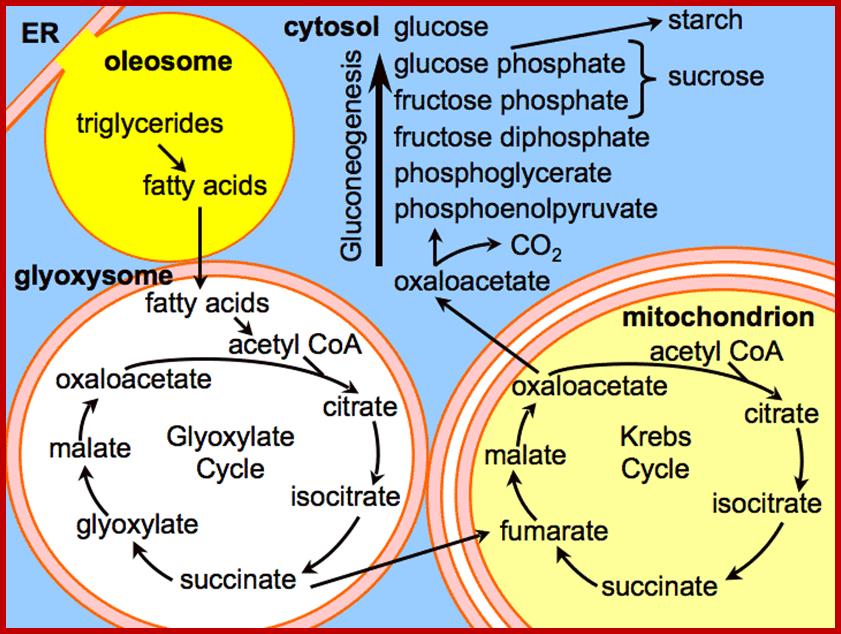 www.look for diagnosis.com; Glyoxylate Cycle
www.look for diagnosis.com; Glyoxylate Cycle
www.lookfordiagnosis.com
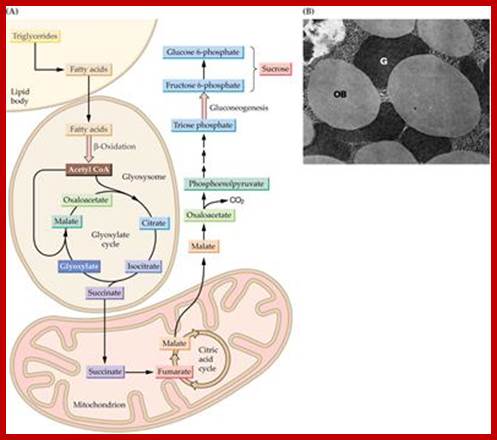
www.imgarcade.com
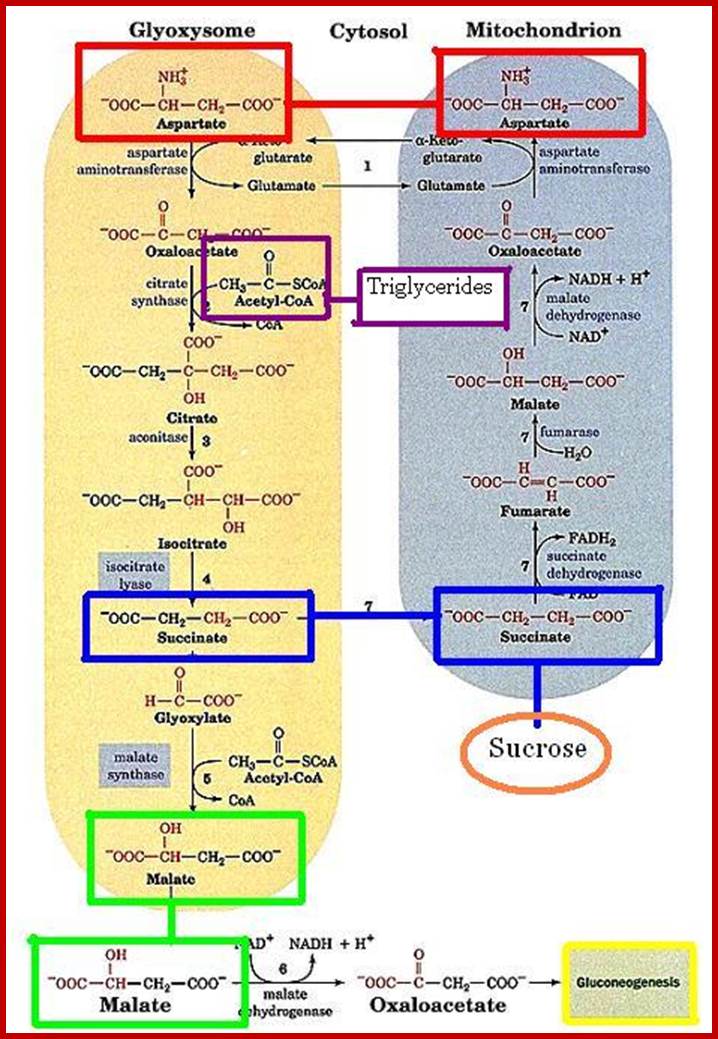
What is the function of glyoxysomes?
What is the function of glyoxysome? Shivago60; http://www.answers.com/
Glyoxysomes (as all peroxisomes) contain enzymes that initiate the breakdown of fatty acids and additionally possess the enzymes to produce intermediate products for the synthesis of sugars by gluconeogenesis. The seedling uses these sugars synthesized from fats until it is mature enough to produce them by photosynthesis Glyoxysomes are specialized peroxisomes found in plants (particularly in the fat storage tissues of germinating seeds) and also in filamentous fungi. As in all peroxisomes, in glyoxysomes the fatty acids are hydrolyzed to acetyl-CoA by peroxisomal β-oxidation enzymes. Besides peroxisomal functions, glyoxysomes possess additionally the key enzymes of glyoxylate cycle (Isocitrate lyase and malate synthase) which accomplish the glyoxylate cycle bypass. This is the most interactive organelle with other cell organelles such as mitochondria, cytoplasm and even chloroplasts.
Biogenesis of glyoxysome is similar to
Peroxisomes, by bit by bit of vesicles carrying the requires components are
fused with the existing underdeveloped pre-glyoxysomes; gradually it enlarges
and become independent organelle.
Functions:
The most important function of glyoxysomes is to convert stored lipids into carbohydrates; which are required for the growing seeding. The conversion of lipids in to glucose requires co-coordinated functions of glyoxysomes, mitochondria and proplastids.
Primarily, glyoxysomes first convert lipids into glycerol and fatty acids; then the latter is subjected beta-oxidation. In germinating seeds, the acetyl CO-A produced during b-oxidation is not drawn into Kreb’s cycle, but it is utilized in glyoxylate cycle. The succinate, produced in glyoxylate reactions, is transported across the membranes into mitochondria where it is further converted into glucose phosphate through phosphoenol pyruvate pathway. The last part of the reaction takes place in proplastids. Besides oxidation and glyoxylate reactions, glyoxysomes also contain urate oxidase and allantoinase responsible for the conversion of urate into allantoin.

Interaction between chloroplasts peroxisomes and mitochondria; http://faculty.ksu.edu.sa/

www.slideshare.net